Home Boris Sidis Archives Table of Contents Next Chapter
|
AN EXPERIMENTAL STUDY OF SLEEP Boris Sidis, Ph.D., M.D. Boston: R. Badger, 1909 |
CHAPTER XII
Cell-energy, Threshold, “Stimulus-exhaustion” and Sleep
WE may advance in a tentative way the following theory which should be regarded as a provisional hypothesis of the causation of sleep.
One of the main characteristics of living protoplasm is its adaptability to the conditions of the, external environment. External stimuli give rise to reactions of adjustment on the part of living substance. This property known in physiology as irritability is specially characteristic of all living matter or of what Huxley so aptly describes as “the physical basis of life.” Verworn defines the irritability of living substance “as its capacity of reacting to changes in its environment by changes in the equilibrium of its matter and its energy.” In other words, living tissue responds to external stimulations with some discharge of energy. The form of the discharge depends on the peculiar protoplasmic structure, according as it is muscle, gland, nerve cell or but slightly differentiated protoplasm, such as amoeba or bacterium. The character of the reaction to stimuli depends on the state of organization of the living tissue. The delicacy of response of living matter to external stimuli has its limit. Very weak stimulations do not call forth any reaction. Living tissue can only be set into activity by stimuli of certain intensity. If the stimulus falls below that intensity, the protoplasm does not react. This holds true of all cells, from the simplest bacterium and infusorium to the most highly differentiated muscle-cell or neuron. The minimal intensity below which the stimulus remains ineffective is regarded as the threshold of stimulation. As Howell puts it: “A stimulus too weak to give a response with a motor nerve is usually designated in physiology as subliminal; a similar stimulus with sensory nerves is frequently expressed by the equivalent term of subliminal, that is below the threshold, so a stimulus just strong enough to provoke a perceptible reaction is the minimal stimulus for efferent nerves and the threshold stimulus for sensory nerves.” “Exceedingly feeble stimuli” says Landois “are without effect. The degree of intensity of stimulation that originates the first trace of sensation is called the threshold of sensation or the threshold-value.” The same is more clearly put by Verworn:
“Let us imagine an organism or part of an organism, e.g., a muscle, under conditions in which no stimulus affects it, and let us bring to bear upon it a stimulus, e.g., the galvanic current, which varies in intensity from zero upward and can be graded easily and delicately. Then we should expect the muscle to exhibit phenomena of stimulation, i.e., to perform a contraction, as soon as the intensity is increased above 0. But this does not happen. The intensity can be increased considerably before the muscle performs even the slightest twitch. Only when the intensity has reached a certain degree does the muscle respond with a contraction; from here on the contraction is never wanting, and up to a certain degree becomes more energetic the more the intensity is increased. The stimulus, therefore, begins to operate only at a certain intensity, and this point is termed the threshold of stimulation. Below the threshold the stimulus is ineffective; above it the effect increases with the increasing intensity of stimulus. For the different forms of living substance the value of the threshold is very different. Thus, nerve-fibres are put into activity by extremely feeble galvanic stimuli, while Amoeba demands very strong currents. The same is true of all other varieties of stimuli in relation to the various forms of living substance.”
Psychologically we may agree with Stout that “the point at which it (the stimulus) is just indistinguishable,—so that the least increase would make it distinguishable is called stimulus-threshold.”
Külpe’s definition is short; “the just noticeable stimulus is technically termed the stimulus-threshold (die Reizschwelle).”
With the increase of stimulations the irritability of the living substance diminishes, the threshold rises. The same minimal stimulus will no longer bring about a reaction, the stimulus must be increased in intensity before any effect can take place. Perhaps the best account of it is given by Verworn in his “General Physiology.”
“If a living object be stimulated by long-continued, oft-repeated, or very strong stimuli, after some time it passes into the condition of fatigue. The general characteristic of fatigue is a gradual decrease of the irritability of the living substance. This is expressed especially in the fact that with increasing fatigue, the intensity of the stimulus remaining the same, the result of the stimulation becomes constantly less.
“We have already become acquainted with some examples of this fact in considering galvanic stimulation. If a constant current of average strength be passed through an Actinosphoerium, at the moment of making, there begin to appear at the anode marked phenomena of contraction. The protoplasm of the pseudopodia flows centripetally until the latter are drawn in. Then the walls of the vacuoles break; and a granular disintegration of the protoplasm results, which proceeds constantly farther from the kathode during the passage of the current. This disintegration, beginning with great energy, becomes slower and less extensive the longer the current flows, and after some time is at a complete standstill. This means that the living substance of the Actinosphoerium becomes fatigued in the course of continual stimulation, and decreases in irritability; hence the stimulus, which at first induced pronounced phenomena of disintegration, later produces no reaction at all. Pelomyxa is fatigued still more rapidly than Actinosphoerium. Stimulation for a few seconds is sufficient to make individuals of this genus wholly non-irritable to currents of equal intensity; a much greater intensity is then required to call out the same reaction.”
The principle of variability of stimulation is of great importance in the reaction of nerve-tissue. When the stimulus remains invariable, both in intensity and quality, no reaction follows. This is clearly brought out in experiments on nerve-tissue. “The electrical current” says Landois,1 “exerts its strongest irritant effects upon a nerve at the time of its entrance into the nerve and at the time of its disappearance. In like manner any rapid increase or decrease of the current passing through a nerve has a strong irritant effect. If on the other hand, the current be allowed to pass gradually into the nerve trunk or to disappear, or the current passing through the nerve be gradually increased or diminished, the visible signs of nerve irritation are much less marked. In general, the stimulation is most pronounced the more rapid the current-variation within the nerve, that is the more suddenly the strength of the current passing through the nerve is increased or diminished.” This holds true in the case of the nerve, in which as Bowditch has shown there is little or no fatigue; where fatigue is present the principle of variability becomes a factor of the utmost consequence. The principle of variability of stimulation plays a very important role in cells in general, and in nerve-cells in particular where fatigue easily sets in and the threshold is raised with the continuation of stimulation and with the successive discharges of cell energy. Variability of stimulation and fatigue influence the fluctuations of thresholds.
It may be well to bring here the recent work of Sherrington in regard to the relation of neuron threshold and the discharge of neuron energy. Sherrington points out that among the characteristic differences between conduction in nerve-trunks and in reflex arcs (where nerve-cells are interpolated), we find in the latter “irreversibility of direction instead of reversibility as in nerve-trunks, fatigability in contrast with the comparative unfatigability of nerve-trunks,” and “much greater variability of the threshold of stimulus than in nerve-trunks.” Sherrington points out that "in certain cases, especially in invertebrata, observation (Apathy, Bethe, etc.), indicates that many nerve cells are actually continuous one with another. It is note-worthy that in several of these cases the irreversibility of direction of conduction which is characteristic of spinal reflex arcs is not demonstrable; thus the nerve net in some cases e.g. Medusa, exhibits reversible conduction (Romanes, Nagel, Bethe and others). But in the neuron-chains of the gray centred system of vertebrates histology on the whole furnishes evidence that a surface of separation does exist between neuron and neuron. And the evidence of Wallerian secondary degeneration is clear in showing that that process observes strictly a boundary between neuron and neuron and does not transgress it. It seems therefore likely that the nexus between neuron and neuron in the reflex arc, at least in the spinal arc of the vertebrae involves a surface of separation between neuron and neuron” “In view therefore of the probable importance physiologically of this mode of nexus between neuron and neuron it is convenient to have a term for it. The term introduced has been synapse.” “At each synapse there is a neuron threshold. At each synapse a small quantity of energy, freed in transmission, acts as a releasing force to a fresh store of energy not along a homogeneous train of conducting material as in a nerve-fibre pure and simple, but across a barrier which whether lower or higher is always to some extent a barrier.”2
Assuming then the principle of variability of stimulation of neuron energy and neuron threshold in regard to the liberation of neuron energy, principles advanced in my previous works, we can return to our subject under discussion, namely sleep. We may regard sleep as a reaction of protoplasm and as such we may express it in terms of neuron threshold and neuron energy liberation.
If a series of stimulations are kept up without variation, the sensory threshold gradually rises and finally the stimuli fall out of consciousness, they fail to awaken the psycho-physiological systems which have responded to the same stimulations before, because of the rise of the thresholds. This psycho-physiological law, characteristic of all tissues and psycho-neural systems, underlies the phenomena of sleep. As the cell or the neuron keeps on reacting to stimulations the disposable physiological energy becomes lowered and there is greater economy in the liberation of cellular or of neuron-energy. The cell or neuron does not respond to the same intensity of stimulus with the same amount of energy. In other words, the threshold rises. Should the stimulation keep on acting without variation, both as to quantity and quality, the threshold rises so high that the stimulus can no longer step over the threshold and can no longer call forth any reaction in the cell. We say that the cell or the neuron is fatigued or exhausted. Really this is not so much due to the exhaustion of the cell as to the exhaustion of the stimulus,—it is the stimulus that has exhausted itself. Vary the stimulus in quantity or in quality and the cell or neuron reacts once more. We may possibly best describe this general physiological fact of cellular reaction by the term of “stimulus-exhaustion.” By repetition the stimulus exhausts itself and can no longer call forth a reaction in the cell, although the cell may otherwise possess a large amount of disposable energy.3
As far as the particular stimulus is concerned the cell may be regarded as asleep. The stimuli are withdrawn. Putting the same general law in different terms we may say that sleep is produced by monotony.
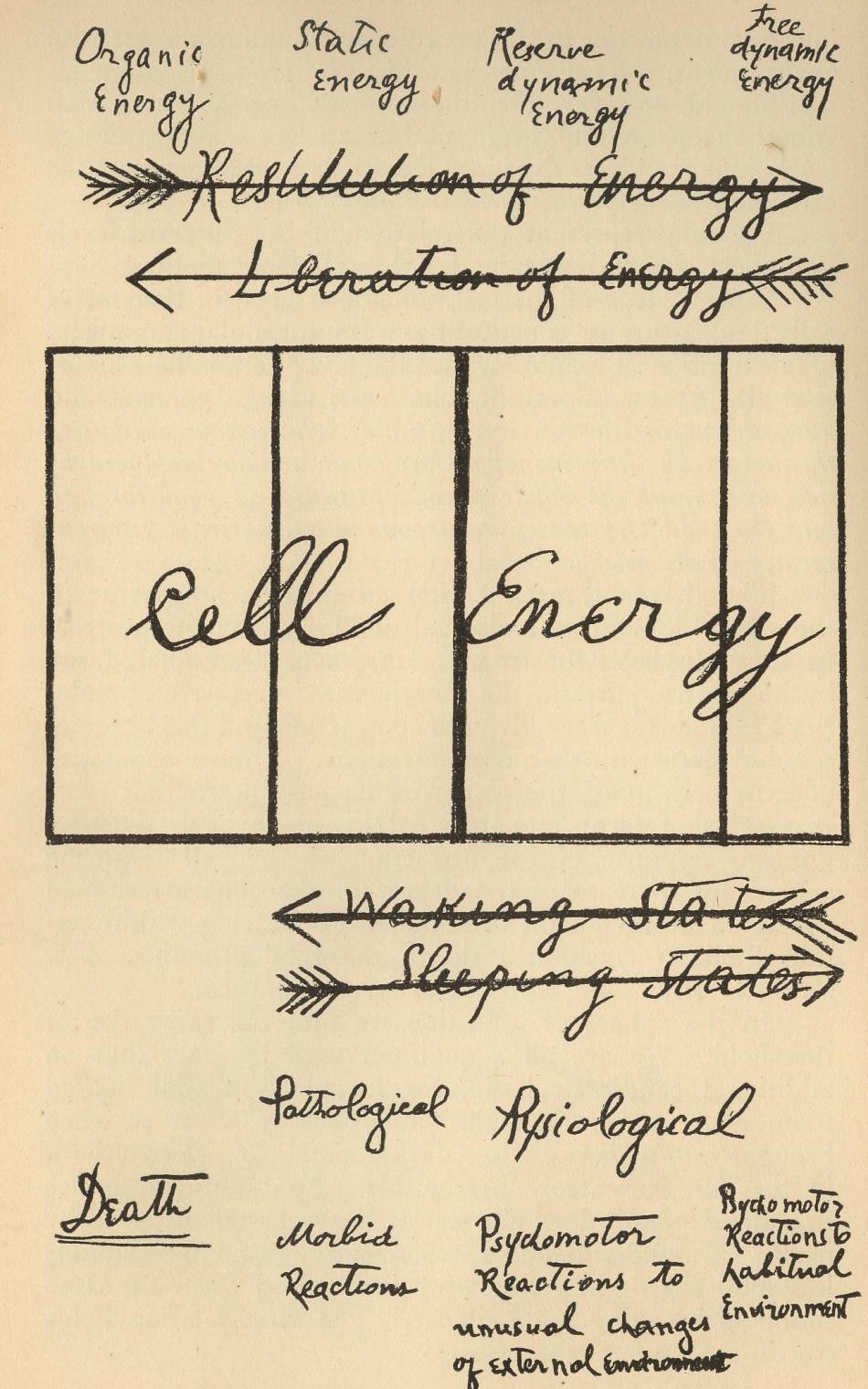
The cell or the neuron may be regarded as a reservoir of energy. With the great biologist, Strassburger, we may describe the cell as “energid.” Conceived in terms of energy the cell possesses various levels of energy. The main levels of cellular energy have been described by me in former works as dynamic, static and organic.
By dynamic energy is meant that portion of energy which the cell can dispose of in its relations and reactions to the stimuli of the external environment.
By static energy is designated that portion of energy which is used only for the life maintenance of the cell, both in relation to other cells and to its own molecular constitution. It is the energy requisite to keep up its morphological organization and internal physiological functions.
By organic energy is meant the energy contained in the tissues of the dead cell not as yet decomposed into its inorganic constituents.
The levels of cellular energy are not different in kind. They merely represent progressive phases or stages of the same process of cellular activity.
In its relations with the external environment, the cell does not utilize the whole of its dynamic energy. A large amount of it lies fallow, so to say, and remains inaccessible to the ordinary stimulations of the external environment. This amount of unused energy may be termed stored, reserve energy.4 For the cell stores up energy as it stores food, in order to be able to meet the various emergencies that may arise in the course of its relations with the external world. In respect to storing up energy the law of stimulus-exhaustion may be regarded as a safeguard to the cell. The same qualitative stimulus cannot draw more than its allotted portion of energy. If the stimulation continues and attempts to draw more than its share the door, so to say, closes and the stimulus knocks in vain against a locked door. The threshold rises with each successive stimulation. When the maximum amount is drawn, there is no longer response to that particular stimulus, or rather to say, to that particular qualitative stimulation of a definite intensity. In regard to it the cell no longer reacts,—it is asleep. In order to respond again to the particular stimulus of a certain intensity and quality, the cell must recuperate its special form of energy. When this recuperation is effected, the cell is once more ready to react to the given stimulus.
The liberation of cellular energy in response to external stimuli may be regarded as the active waking states, while the restitution or passive rest-states of the cell, during which the threshold is raised and cellular recuperation takes place, may be regarded as sleeping states.
From the standpoint of cell metabolism, waking states are correlative with katabolic changes, while sleeping states are correlative with anabolic conditions. When the cell is active and awake, it liberates energy; when it sleeps, it stores energy. This process of storing up energy is going on all the time to some extent. If a stimulus of a certain intensity and quality keeps on liberating energy, it may arrive after some time at the maximum, when the threshold rises so high that the cell no longer reacts.5 In respect to that stimulus the katabolic, active, waking state ceases and is replaced by the anabolic, sleeping states. The cell may thus be awake to all other stimulations, but in regard to the special stimulus it is asleep. We may say that the cell is asleep partially. When the liberation of energy has reached the maximum point, and the thresholds are raised in regard to most or to all the ordinary stimuli of the external environment, the cell may be said to be fully asleep.
Should however, the liberation of energy by various stimuli keep on and begin to liberate the reserve energy levels and then pass over into the levels of static energy, the cell falls into a pathological waking state.
We may represent the relation of the different levels and their correlative states by the following diagram. What is true of the cell holds also good of systems of cells and neurons constituting a multicellular organism. The condition of monotony and the law of stimulus-exhaustion play a very important role in the various changes and adaptations of life-activity. Stimuli which have exhausted themselves by their monotony drop out and are replaced by new ones until the whole round of stimuli is gone through and the organism ceases to respond to its external environment,—falls asleep.
The rise of threshold after stimulation holds true in the whole domain of biological activities. If the gastrocnemius muscle of the frog, for instance, is stimulated, say by an electric current, the muscle with successive stimulations responds less readily with contraction and this becomes more evident with the onset of fatigue. Pffefer in a series of extremely interesting experiments has shown that spermatozoids of ferns are attracted by malic acid, the progressive response of the attraction of the cell requiring a constant increase of the degree of the concentration of the acid, the increment of stimulations bearing within certain limits a constant ratio to the total stimulus. The threshold rises with each successive stimulation.
In the sphere of sensation we find the same rise of threshold. We are all acquainted with the fact that an additional candle or lamp, for instance, in well lighted room does not produce the same sensory effect as when brought into a more or less dark room. An electric light in the sun is scarcely perceptible. An additional ounce to a lifted pound does not feel as heavy as when raised by itself. A sound added to another sound or to a noise sounds less loud than when appearing isolated or when the same sound is breaking upon silence. The same relation holds true in the case of other senses.
That same truth is still more clearly brought out in the fact that if we take a certain stimulus as a unit giving rise to a definite sensation, then as we progressively ascend and add more and more units of the same stimulus, the qualitative intensity of the sensation is far from rising proportionately. If we take for instance, the weight of an ounce as our unit of stimulation, then the successive moments of unit-stimulations, that is of ounces, will not give rise to as distinct and similar sensations as did the initial stimulation. The second ounce will give a sensation fainter than the first one and the third fainter than the second and so on until a point is reached, when the sensation of an additional ounce will not at all be appreciated, will dwindle away and almost reach the zero-point. In the same way, if the pressure of a gramme is excited in the hand, successive increments of grammes will not in equal degree increase the sensory effect. The additional increments of grammes, though they are equal units of stimulation, give rise to fainter and fainter sensations, until finally all sensory appreciation of the added unit fades away and disappears. If the hand is immersed in water say at the freezing point, an addition of ten degrees will be perceptibly appreciated, while successive increments of ten degrees each will be felt less and less and finally will not be noticed and will become difficult to detect. In short, the threshold rises with each successive stimulation.
To bring about a sensory response of an already stimulated sense-organ the intensity of the stimulus must be relatively increased. This is what constitutes Weber’s law. The continuous progressive sensory response of a sense-organ requires a constant increase of stimulation which within certain limits, bears a constant ratio to the total stimulus.6 This law is sometimes summed up by physiologists and psychologists in the statement that the stimulus rises in a geometrical and corresponding sensation in an arithmetical progression, or as Fechner expresses it, “the sensation varies as the logarithm of the stimulus.” We are probably nearer the truth, if we limit ourselves to the statement that “the increase of the stimulus necessary to produce a just perceptible difference in the sensation bears a constant ratio to the total stimulus.” In short, activity raises the threshold. The phenomena fall under the law of stimulus-exhaustion. Each reaction to an external stimulus raises the threshold of that stimulus and with its repetition becomes finally excluded, temporarily, at least, from the cycle of living stimuli, that is, such as are capable of bringing about reactions and adjustments to the external environment. The excluded stimulus is ignored, becomes non-existent for the organism. The organism is asleep for that stimulus.
In the course of the daily activity of the individual the thresholds of sensitivity and of sensori-motor reactions in response to the stimuli of ordinary life rise gradually, the stimuli fall below the threshold of living reactions. The mass of stimuli of the external world constituting the external environment of the individual drops out of the life-cycle of the individual and for the time being ceases to exist for him. The individual is asleep, asleep for that environment. In other words the stimuli of the external world by their continuous action have become monotonous, have raised the thresholds and have become excluded from the life existence of the individual who is now deeply asleep for that given environment.
Should an appreciable variation occur in the environment at about the time when the organism is ready to fall asleep the external world becomes again a living reality. The individual becomes alive and wide awake to what is going on around him. If an appreciable change in the total mass or in some stimuli constituting the environment takes place, the individual may be awakened out of his sleep. Should the variation occur in some of the stimuli, their thresholds become lowered and the individual, though asleep in regard to all else, is awake in regard to them. This partial waking state is the soil in which dreams develop most luxuriantly. We do not dream, when we are asleep, we dream, when we are awake.7
The individual may be asleep to all else and still be awake to special stimuli whose thresholds are very low. The mother watching over her babe, the nurse, the physician attending their patients may be sound asleep to loud noises, but are alive and wide awake to the slightest changes in their charges. They have lost touch with the whole external world, but the ones cared for still have a firm hold on them. High as our bridges are raised for the whole world, they are low for the ones we care for.
Let an appreciable change occur in the stimuli to which we are awake, an amount of energy immediately is drawn from the reserve store, the thresholds barring the entrance to the host of intruders are lowered and we are once more in communication with the external world,—our sleep is gone and we are wide awake. We can thus be wide awake in the very depth of our sleep. The possibility of shaking off the grip of sleep under appropriate circumstances and especially the highly significant fact of wakefulness in the very depths of sleep form insuperable difficulties for all those plausible, apparently scientific theories of sleep, theories based on circulation, engorgement, anemia or hyperaemia, narcosis and autointoxication. We can watch in our sleep, count the flow of time and awake at the right moment. The fact that we can sleep and still watch and keep awake in regard to special objects and particular persons shows that sleep is not a matter of blood circulation or of intoxication by accumulation of waste products in the system. Sleep is not a disease, not a pathological process due to the accumulation of toxic products in the brain or in the system generally. Sleep is not of those fatal uncontrollable onsets characteristic of the morbid processes, such as epilepsy, sunstroke, or apoplexy, nor is sleep a sort of narcolepsy. Sleep is not a disease, it is not, as the chemical speculators would have it, a kind of narcosis of the system by the poisons of fatigue products. Sleep is not pathological, it is essentially physiological in character. We do not go to sleep, because we are poisoned and wake up when the poisons are eliminated. We go to sleep at our own sweet will and can wake at pleasure. We can wake at any moment and can even be awake in the very depths of our sleep. Sleep is not an abnormal condition, it is a normal state. Like the waking states, sleep-states are part and parcel of the life-existence of the individual. Waking and sleeping are intimately related,—they are two different manifestations of one and the same life-process,—one is as normal and healthy as the other. One cannot help agreeing with Claperède’s biological view that sleep is a positive function of the organism, that sleep belongs to the fundamental instincts. As Claperède forcibly puts it: “Le sommeil est une fonction de defense, un instinct qui a pour but, en frappant l’animal d’inertie, de l’empêcher de parvenir au stade d’epuisement: ce n’est pas parce que nous sommes intoxiqués, ou épuisés, que nous dormons, mais nous dormons pour ne pas l’être.”
If we look at the matter from a psychological standpoint, we may say that sleep is a rise of the thresholds of mental aggregates or of moments consciousness. I have pointed out in a former work8 of mine that to minimize the expenditure of neuron energy and reach the minimum of consciousness constitutes the tendency of psychomotor life.
We can fully realize the importance of this tendency, if we regard it from a teleological point of view. In the struggle for existence or in the economical system of competition of modern life the saving of unnecessary expenditure, where only possible, is of the highest consequence. Organisms that can best effect such an economy of energy will be best fitted to survive. Those organisms that are enabled to reduce to its minimum the friction and loss of neuron energy have the advantage of possessing at their disposal a greater amount of energy to cope with new circumstances, with novel conditions, and react better and in a more favorable way when confronted with changes in their environment. This economizing becomes absolutely indispensable in the life-existence of higher organisms, the environment of which is highly complex. The reduction of psychomotor activity to the least amount of psychophysiological expenditure, in other words, to the minimum of consciousness is the law of psychomotor life in general and of the highest representatives of that life in particular.
I have further pointed out in the same work that this economizing is by no means an endless process, there is a certain minimum of consciousness beyond which psychic states cannot pass. This minimum of consciousness once reached, must remain stationary, for a fall below it is the arrest of the activity of the mental aggregate. In other words, there is a certain minimum below which consciousness cannot be reduced with impunity. Reduce the consciousness of the total psychic aggregate by lowering the sensibility of its constituents and the whole mental system will cease to function. Now under the conditions of monotony and limitation of psychomotor activity the moment-threshold rises until the psychic minimum vanishes and the organism is asleep.
Putting the matter again in teleological terms we may say that we go to sleep when we relinquish our hold on the relations of our external environment. We fall asleep when our consciousness is fagged, when we wish no longer to enter into communication with the external world, when we lose interest in our surroundings. When our interest in external existence fags and fades away, we go to sleep. When our interests in the external world cease, we draw up the bridges, so to say, interrupt all external communication, as far as it is possible, and become isolated in our own fortress and repair to our own world of organic activity and inner dream life. We fall asleep when the vital interests in external being have fallen into the background of consciousness; we awake when those interests are aroused. When the struggle for existence ceases we repair to our castle and battlements. Sleep is the interruption of our intercourse with the external world; it is the laying down of our arms for a respite in the struggle of life. Sleep is a truce with the world. When all psychomotor reactions to the stimuli of the external environment cease, we sleep. We sleep, because we are no longer interested to take an active part in the battle of life. From a teleological standpoint we may say that sleep is a dismissal of the external world with all its vicissitudes, troubles and pains. We cease to desire, we cease to react, and we sleep and dream in peace.
|
And when evening descended from heaven above, And the earth was all rest, and the air was all love, And delight, tho’ less bright, was far more deep And the day’s veil fell from the world of sleep, And the beasts, and the birds, and the insects were drowned In an ocean of dreams without a sound; Whose waves never mark, tho’ they ever impress The light sand which paves it, consciousness. —Shelly. |
_____________
1. Text-book of Physiology.
2. Sherrington, “The
Integrative Action of the Nervous System.” This may be related to the
interesting experiments referred to by Sherrington:
“When the scratch-reflex elicited from a spot of skin is fatigued, the fatigue
holds for that spot, but does not for the reflex as obtained from the
surrounding skin. The reflex is then tired out to stimuli at that spot easily
obtainable by stimulation two or more centimetres (half an inch) away. This is
seen with either mechanical or electrical stimuli.”
(The Integrative Action of the Nervous System, Sherrington)
3. “The singleness of
action from moment to moment thus assured is a keystone in the construction of
the individual whose unity it is the specific office of the nervous system to
perfect. Releasing forces acting on the brain from moment to moment shut out
from activity whole regions of the nervous system, as they conversely call vast
other regions into play.”
“An arc
under long continuous stimulation of its receptor tends even when it holds the
common path, to retain its hold less well. Other arcs can then more readily
dispossess it. A stimulus to a fresh arc has, in virtue of its mere freshness, a
better chance of capturing the common path.”
“This waning
of a reflex under long-maintained excitation is one of the many phenomena that
pass in physiology under the name ‘fatigue.’ Its place of incidence lies at the
synapse. It seems a process elaborated and preserved in the selective evolution
of the neural machinery. It prevents long continuous possession of a common path
by any one reflex of considerable intensity. It favours the receptors taking
turn about. It helps to ensure serial variety of reaction. The organism, to be
successful in a million-sided environment, must in its reactions be many-sided.”
(Sherrington, Address British Association for the Advancement of Science, 1904).
4. See my Studies in
Psychopathology, Boston Medical and Surgical Journal, 1907.
5. “Whenever by a stimulus
applied to an irritable substance, the potential energy there stored up is
liberated the following phenomena may be observed: (1) A so-called latent period
of variable duration during which no effects of stimulation are manifest; (2) A
very brief period during which the effect of stimulation reaches a maximum; (3)
A period of continued stimulation during which the effect diminishes in
consequence of the using up of the substance containing the potential
energy—i.e., a period of fatigue; (4) A period after the stimulation has ceased
in which the effect slowly passes away.
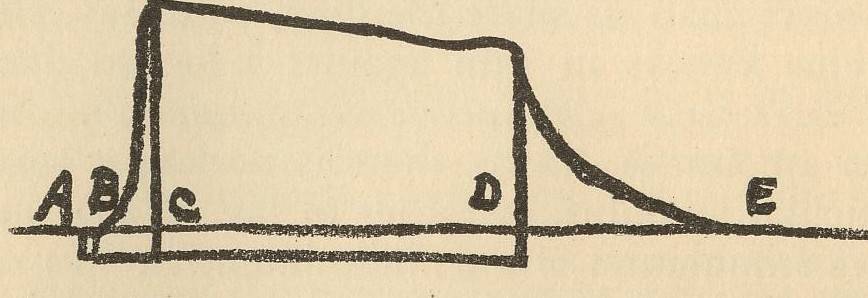
The curve
drawn by a muscle in tetanic contraction, as shown in the diagram, illustrates
this phenomenon. Thus, if AD represents the duration of the stimulation, AB
indicates the latent period, BC the period of contraction, CD the period of
fatigue under stimulation, and DE the after-effect of stimulation showing itself
as a slow relaxation. When light falls upon the retina corresponding phenomena
are to be observed.” An American Text-Book of
Physiology, William H. Howell, Vol. II, p. 343.
6. By noting for each
strength of stimulus the addition required to evoke a just perceptible
alteration of sensation, a series of quantities is obtained expressing the law
according to which sensation alters when stimulation is increased. This
expression is the so-called “law” of Weber. It says that a given stimulus is
perceived less when added to a large than to a small one, or that an addition to
a large stimulus is perceived less than an addition to a small one, unless it,
relatively to the stimulus, is as great. The “law” may be phrased variously in
physiological theory. It may be interpreted as not physiological at all, but
psychological. The disproportion between increment of stimulus and increment of
sensation may take place in purely psychological events and processes. Wundt is
of that opinion. He points to the wide occurrence of such a ratio in all
psychical activity as outcome of the relativity inherent in every conscious
process. Waller finds the response in a nerve trunk directly stimulated, as
judged by action current, increase much more nearly directly as the increase of
external stimulus than does the response from muscle when nerve is stimulated,
or from nerve when retina is adequately stimulated. Waller's evidence seems to
point to the law being in part a function of the nerve-cell endings; probably,
therefore, applicable to synapses as to motor plates. Delboeuf considers the law
an expression of ever-increasing proportion of loss of effect in the central
nervous system, due to “fatigue.” (Sherrington in Schäfer’s Text-Book of
Physiology, Vol. II, p. 931.)
7. My work on dreams will be
published in a separate essay. Dreams
8.
Multiple Personality.
Boris Sidis Archives Contents Next
![]()