| Home
Boris
Sidis Archives Menu Table of Contents
Next Chapter
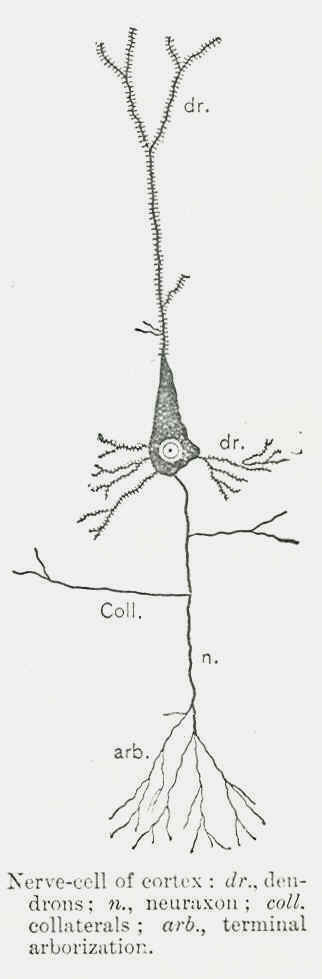
CHAPTER XXI THE PHYSIOLOGY AND PATHOLOGY OF SUBCONSCIOUSNESS THE mental processes of association and aggregation of psychic contents in the synthesis of momentconsciousness and the including of the moments-consciousness in synthesis of higher and higher unities can be expressed in physiological terms of cellular activity. The structure of the cell and its morphological relation to other cells can give us a glimpse into the physiological processes that run parallel to mental synthesis and dissociation. The nerve-cell, as the reader knows, is a nucleated mass of protoplasm highly complicated in its structure and organization. The nerve-cell possesses many filaments or "processes," all of which, called dendrons, branch repeatedly and terminate in a network of multitudes of fibre-processes representing a greater volume than the cell body itself, with the exception of a single process termed neuraxon, which remains comparatively unchanged in its diameter along its whole course and sends out but a few branches called collaterals. The terminals of collaterals and neuro-axons are in their turn split into a comparatively small number of branches called the terminal arborization. If we inquire as to the connection of nerve-cells with one another, we find that no nerve-cell is anatomically connected with other cells. Every nerve-cell with all its processes forms a distinct and isolated morphological individual. Every nerve-cell anatomically considered is a complete unit. The processes coming out from different nerve-cells do not fuse with processes coming out from other nerve-cells, but rather interlace and come in contact, like the electrodes of a battery in forming the electric circuit. Thus neurological investigations point to the highly significant fact that the connections among the nerve-cells are not of an anatomical but of a physiological nature. The association of nerve-cells is not organic, but functional.
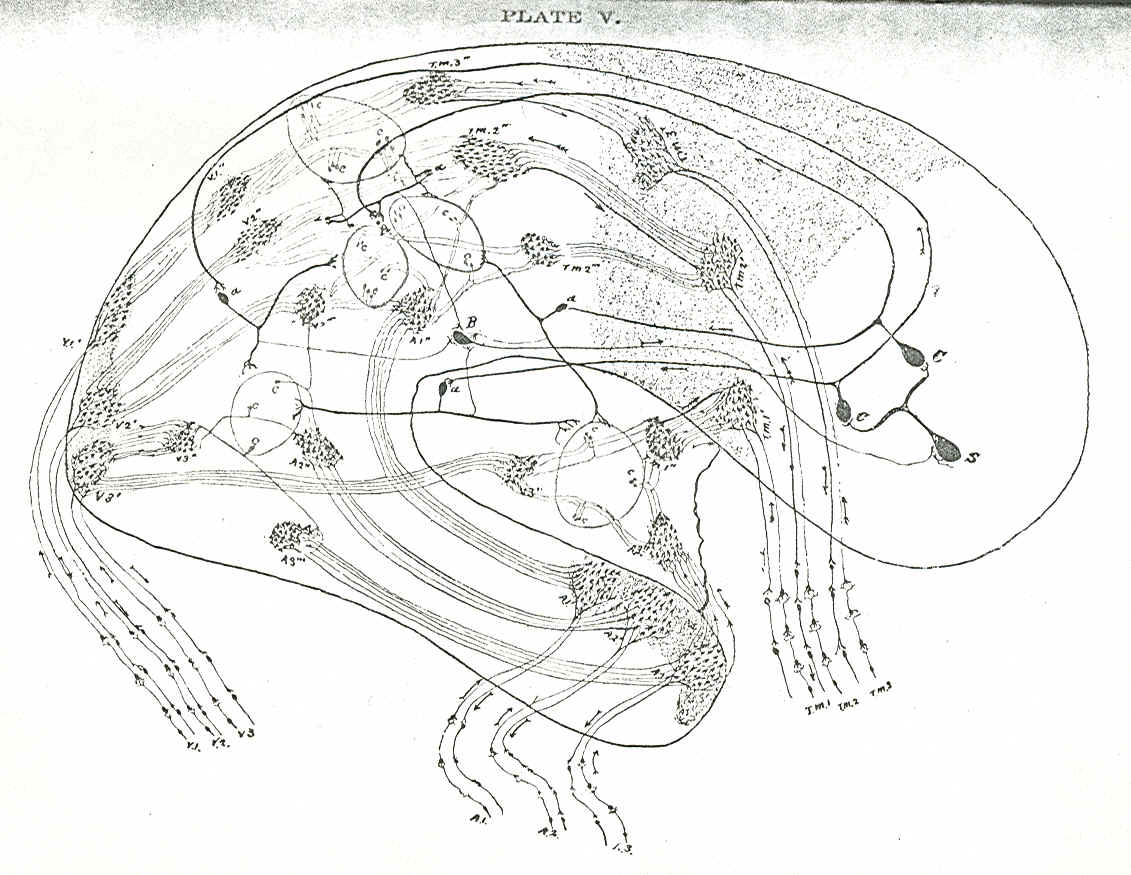
Nerve-cells with concomitant psychic moments-content come into contact with other nerve cells accompanied by psychic content by means of their fine terminal processes. This association of cells forms a group whose physiological function has a concomitant mental activity resulting in some form of psychic synthesis. By means of association fibres the groups arc organized into systems, the systems into communities, the communities into clusters, the clusters into constellations, and each of the higher, more complex aggregates is more feebly organized by less stable association fibres. The combination of groups into systems and of these systems into clusters and constellations by means of association fibres have as their psychic concomitants higher and higher forms of mental syntheses.1 Thus moments-content are synthetized in the unity of moments-consciousness, and the latter are synthetized in their turn in higher and higher unities. The simpler, the less complicated a group of nerve cells is, and the longer and more frequent their fine processes come in contact, the greater is the tendency of that group to form permanent relations; awl the same holds true of system of cells in communities, clusters, and constellations. We may therefore say that the organization of a system or constellation of cells is in proportion to the duration and frequency of their associative activity. Groups of nerve-cells with a more or less stable function become gradually organized and form a sable organization. The more complex, however, a system of nerve is, the greater is its instability, and in the very highest systems or constellations of clusters the instability reaches its maximum. The instability of a system is in proportion to its complexity. In the very highest constellations the instability is extreme, and there is going on a continuous process of variation. Under the action of the slightest external or internal stimuli, such unstable systems or constellations lose their equilibrium, dissolve and form new systems, or enter into combination with other constellations, On the psychical side we have the continuous fluctuation of the content of attention. The characteristic trait of the highest type of psycho-physical life under the ordinary stimuli of the environment is a continuous process of association and dissociation of constellations. As the stimuli increase in their intensity, be they of an external or internal nature―be they toxic, such as the influence of a poison, or purely mechanical, such as the action of a blow, or be they of a pure1y internal psycho-physiological character, such as a strong emotion-a process of dissolution sets in, and the highest, the most unstable, the least organized constellations of clusters are the first to dissolve, With the further increase of the intensity of the stimulus the dissolution goes deeper and extends further―the simpler, the more stable, the more organized systems become dissolved, The psycho-physical content, however, does not disappear with the dissolution of the system; the content exists in the less complex forms of cell-associations, and psychically in the simpler forms of mental synthesis. The same result may be effected by stimuli of less intensity but of longer duration. A durable hurtful stimulus is in fact by far the more detrimental to the life of cell-aggregation. The pathological process of dissociation and disaggregation may be regarded as a function of two factors―of duration and intensity. Such a dissociation is not of an organic but of a functional character. The association fibres that connect groups into systems, communities, clusters, constellations contract. The fine processes of the nerve-cells, the dendrons, or the collaterals that touch these dendrons, thus forming the elementary group, retract and cease to come in contact.2 Association fibres combining the highest constellations are the first to give way; they are the latest to arise in the course of psycho-physical evolution, they are the most unstable, the least organized, and are also the first to succumb to the process of dissolution. The instability of association fibres is proportionate to the complexity and instability if the joined clusters and constellations. At the first onslaught of inimical stimuli the cellcommunities combined into clusters and constellations by association fibres become dissociated and independent of one another. Cell-communities, being more firmly organized than clusters and constellations, of which they are a part, and acting as a more organized whole, resist longer the action of hurtful stimuli. The association-cells that connect different clustered cell-communities contract or retract their fine terminal processes, and the cluster is dissolved. As the hurtful stimuli become more intense, the systems within the cell-community, though more firmly organized by association-fibres than the clusters, withdraw in their turn from the action of the hurtful stimuli. The associationcells that combine systems into communities retract their terminal processes, and the result is the dissolution of the cell-community into its constituent systems, which have more power of resistance than communities of cells, because systems are far more stable, far better organized. As the stimuli rise in intensity the process of disaggregation reaches the systems and they fall asunder into groups. With the further increase of the intensity of the hurtful stimuli the process of disaggregation affects the group itself, the fine processes of the nerve-cell, the dendrons or collaterals and the terminal arborization of the neuraxon contract, withdraw from the hurtful stimuli, as the monocellular organism retracts its pseudopodia from the influence of noxious stimuli. Thus the groups themselves become dissociated, and are dissolved into a number of simple and isolated nerve-cells. For plan of the organization of brain-cells, see Plate V.
The following experiment, made at my request by Mr. R. Floyd, at the Pathological Institute of the New York State Hospitals, tends to confirm the theory of retractility of the extensions of the ganglion cell protoplasm.
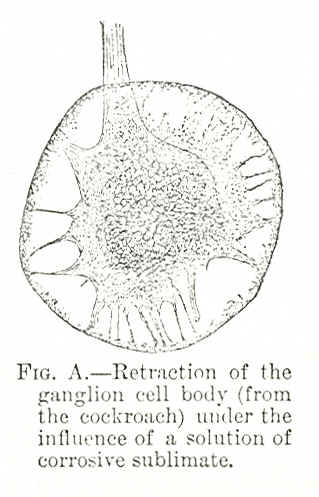
Fig. A shows the retraction of one of the ganglion cells of the cockroach in the living state (Blatta orientalis) under the influence of a strong toxic reagent, corrosive sublimate. The outer circular zone indicates the normal volume of the cell in the living condition, and the retracted outline of the cell indicates the reduction of the volume after contact with the corrosive sublimate. The protoplasmic network of the cell having become contracted under the influence of this toxic reagent, the inference seems to be presented that the fibrillæ of the dendrons, and perhaps of the axon also, which are continuous with the fibrillar network in the cell-body, may become correspondingly retracted. The dendrons are not shown in the preparation, but the root of the axon with its parallel fibrils continuous with the cell-body network is shown at the right-hand side. This whole process of dissolution is functional, for the disaggregation occurs only in the different forms of cell combinations. The cell itself, however, with all its processes remains intact and organically sound. With the removal, therefore, of the hurtful stimuli, there is once more a tendency, on account of the habit acquired from previous combination, to form old associations, and the old relations and functions are gradually restored. In short, until the process of dissolution reaches the individual cell, the process is not of an organic but of a functional character. All functional diseases are cases of psycho-physiological disaggregation, and the gravity of the disease is proportional to the amount of dissociation. A functional disease or functional change is a disaggregation of clusters and systems of nerve-cells with their concomitant moments-consciousness and moments-contents. This disaggregation consists in the withdrawal of the simpler and better organized cell-colonies from the more complex systems, and, lastly, in the withdrawal of individual cells from the group or cell-colony. The whole process of dissociation or disaggregation is one of contraction, of shrinkage, from the influence of hurtful stimuli. First, the most unstable association-fibres are loosened, and communication is interrupted in the clusters forming the highest and most complex constellations, and then, as the intensity of the stimuli increases, the more stable association-fibres are loosened from the systems they connect. With the further increase of the stimuli the process of disaggregation descends still lower, to the elementary group formed of individual cells; the cells withdraw the terminal processes by which they come in contact with those of other cells in the same group. In post-hypnotic states, in cases that go under the name of hysteria, in many forms of aphasia, in many obscure mental diseases, in many psychic states subsequent to great mental shocks, in many mental maladies known as the "psychic equivalent of epilepsy,"3 we meet with cases of different degrees of cell-disaggregations, accompanied by all shades and forms of mental dissociation or amnesia, forms and types which I shall discuss further on. These forms may be spontaneous, as in cases of diseases, or they may be artificial, as in the case of hypnosis. One psycho-pathological process, however, underlies all the various forms of functional diseases, and that is the process of cell-disaggregation, with its concomitant dissociation of moments-consciousness.4
________________ 1. The
difficulties of how a conglomeration of objective units can possibly give rise
to a unity in a synthesis are excellently well discussed by Prof. W. James in
the first volume of his Psychology. We take it as a postulate that the very
nature of mental activity is synthesis.
|
 A
PLAN OF THE ORGANIZATION OF THE BRAIN CELLS INTO
GROUPS, SYSTEMS, COMMUNITIES, CLUSTERS, AND CONSTELLATIONS
A
PLAN OF THE ORGANIZATION OF THE BRAIN CELLS INTO
GROUPS, SYSTEMS, COMMUNITIES, CLUSTERS, AND CONSTELLATIONS